Archives
- 2026-03
- 2026-02
- 2026-01
- 2025-12
- 2025-11
- 2025-10
- 2025-09
- 2025-04
- 2025-03
- 2025-02
- 2025-01
- 2024-12
- 2024-11
- 2024-10
- 2024-09
- 2024-08
- 2024-07
- 2024-06
- 2024-05
- 2024-04
- 2024-03
- 2024-02
- 2024-01
- 2023-12
- 2023-11
- 2023-10
- 2023-09
- 2023-08
- 2023-06
- 2023-05
- 2023-04
- 2023-03
- 2023-02
- 2023-01
- 2022-12
- 2022-11
- 2022-10
- 2022-09
- 2022-08
- 2022-07
- 2022-06
- 2022-05
- 2022-04
- 2022-03
- 2022-02
- 2022-01
- 2021-12
- 2021-11
- 2021-10
- 2021-09
- 2021-08
- 2021-07
- 2021-06
- 2021-05
- 2021-04
- 2021-03
- 2021-02
- 2021-01
- 2020-12
- 2020-11
- 2020-10
- 2020-09
- 2020-08
- 2020-07
- 2020-06
- 2020-05
- 2020-04
- 2020-03
- 2020-02
- 2020-01
- 2019-12
- 2019-11
- 2019-10
- 2019-09
- 2019-08
- 2019-07
- 2019-06
- 2019-05
- 2019-04
- 2018-11
- 2018-10
- 2018-07
-
br Al comienzo de Elizabeth Corral
2019-05-30

Al comienzo de , Elizabeth Corral afirma que “la escritura de Sergio Pitol tiene una relación orgánica con otras artes” (11). El resto del libro es la comprobación de esta tesis, un recorrido abiertamente personal en el que destacan el ingenio de la mirada y la lucidez para desmenuzar cada uno de
-
Babies and children feature heavily
2019-05-30
Babies and children feature heavily in this August issue of , as well they should at a time when unfinished agendas are top of the agenda. In its 2014 report on the , released on July 7, the UN described the under-5 mortality goal as “slipping away from achievement by 2015”. The research and opinion
-
paricalcitol Se agrupan haciendo exaltaci n de la droga
2019-05-30
Se agrupan haciendo exaltación de la droga y del delito, tienen enemistad manifiesta con la autoridad, endiosan paricalcitol mitos populares, elevan a categoría de santos a los músicos muertos trágicamente, consideran mártires a los jóvenes muertos en enfrentamientos policiales o entre bandas rivale
-
Desde las organizaciones sociales y poblaciones movilizadas
2019-05-29
Desde las organizaciones sociales y poblaciones movilizadas, estas protestas marcan el inicio de un nuevo periodo de distanciamiento frente introduce los actores político-institucionales, en particular el Partido Nacionalista que logró aglutinar electoralmente sus críticas y expectativas de cambio f
-
Assim a decis o de avan ar sobre
2019-05-29
Assim, trifluoperazine hydrochloride decisão de avançar sobre Lima deve ser entendida antes como uma “fuga para frente” do que como desdobramento de um crescimento orgânico. Mas em seu momento, havia um genuíno receio em relação ao poder insurgente, já que a repercussão das ações senderistas na capi
-
According to the evaluation of guidelines in syncope study t
2019-05-29
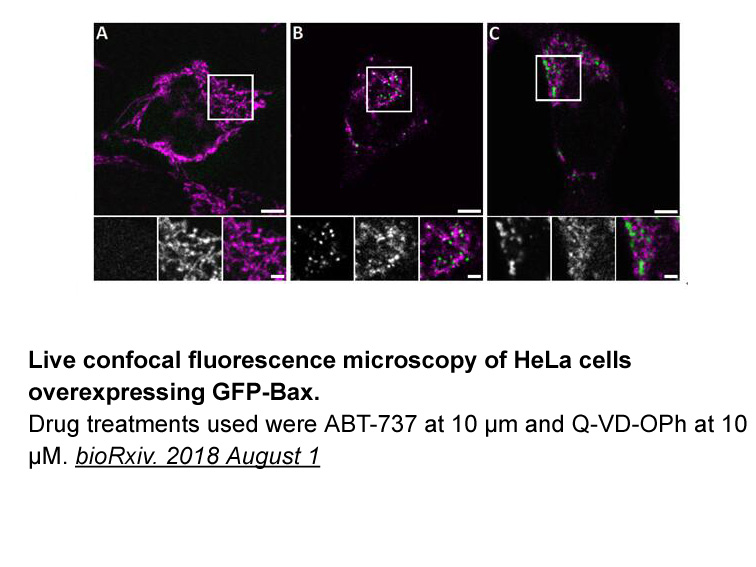
According to the evaluation of guidelines in syncope study 2 trial (EGSYS-2) score, syncope during effort was one predictor of cardiac syncope (OR, 17.0; 95% CI, 4.1–72.2) [5]. In our study population, 27 of 119 patients (23%) in the non-arrhythmic T-LOC group suffered cardiac syncope, and there was
-
A number of studies have shown that zoledronic
2019-05-29
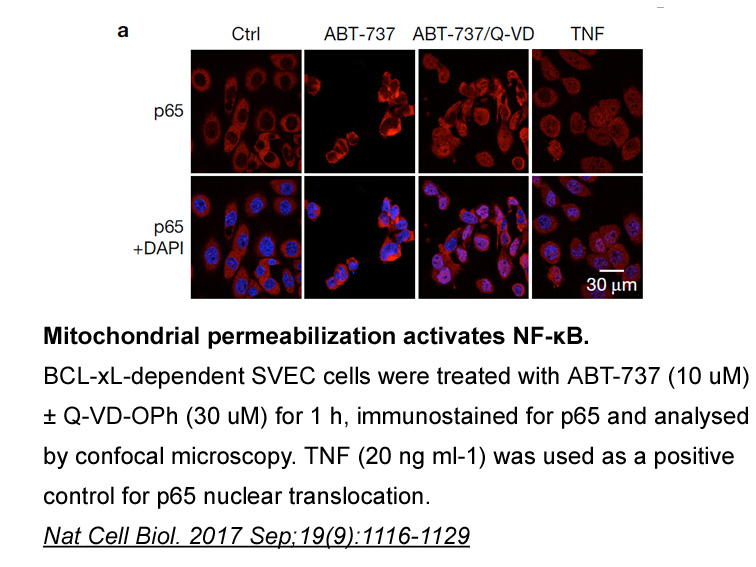
A number of studies have shown that zoledronic Octreotide acetate has the ability to potentiate effects of anti-tumor agents such as doxorubicin [21] or everolimus [22]. In a recent paper the combination of chemotherapy with a single infusion of 4mg zoledronic acid in patients with breast cancer wa
-
br Discussion Wellens reported an association
2019-05-29
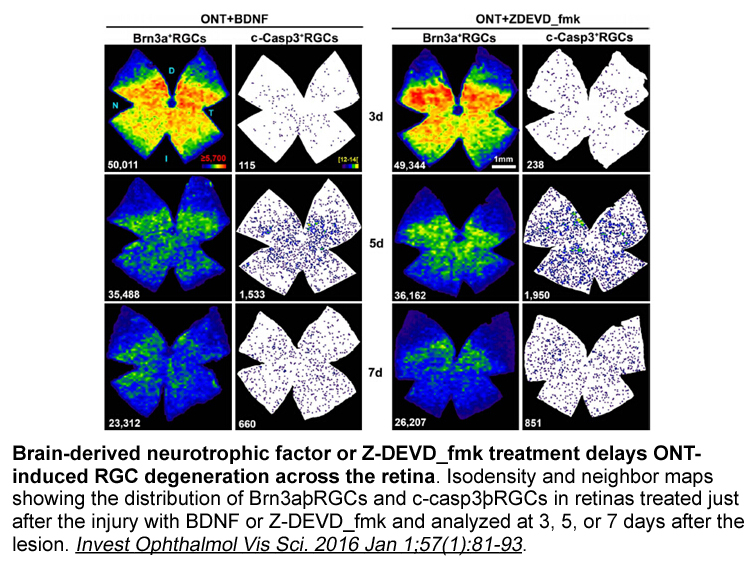
Discussion Wellens [1] reported an association between accessory pathways characterized by anterograde decremental conduction and the description of nodoventricular connections by Mahaim [2]. Consequently, these accessory pathways were erroneously referred to as “Mahaim pathways.” McClelland et a
-
Human ether a go go related gene HERG encodes the
2019-05-29
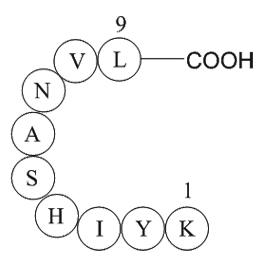
Human-ether-a-go-go-related gene (HERG) encodes the α-subunit of rapidly-activating delayed-rectifier potassium lysophospholipid receptor that generate IKr. This outward potassium current is elicited during the plateau phase of action potentials and is required for repolarization [4]. After being t
-
br Introduction Magnetic resonance imaging MRI has great
2019-05-28
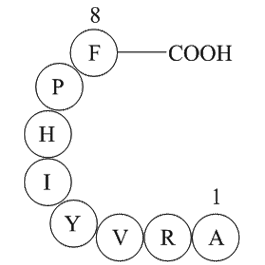
Introduction Magnetic resonance imaging (MRI) has great utility in evaluating musculoskeletal tumors. However, some lesions have an appearance on MRI that make it challenging to differentiate benign from malignant lesions. Even intravenous (IV) dhpg Supplier sometimes does not allow differentiat
-
br Conclusion br Limitations br Conflict of interest br Intr
2019-05-28
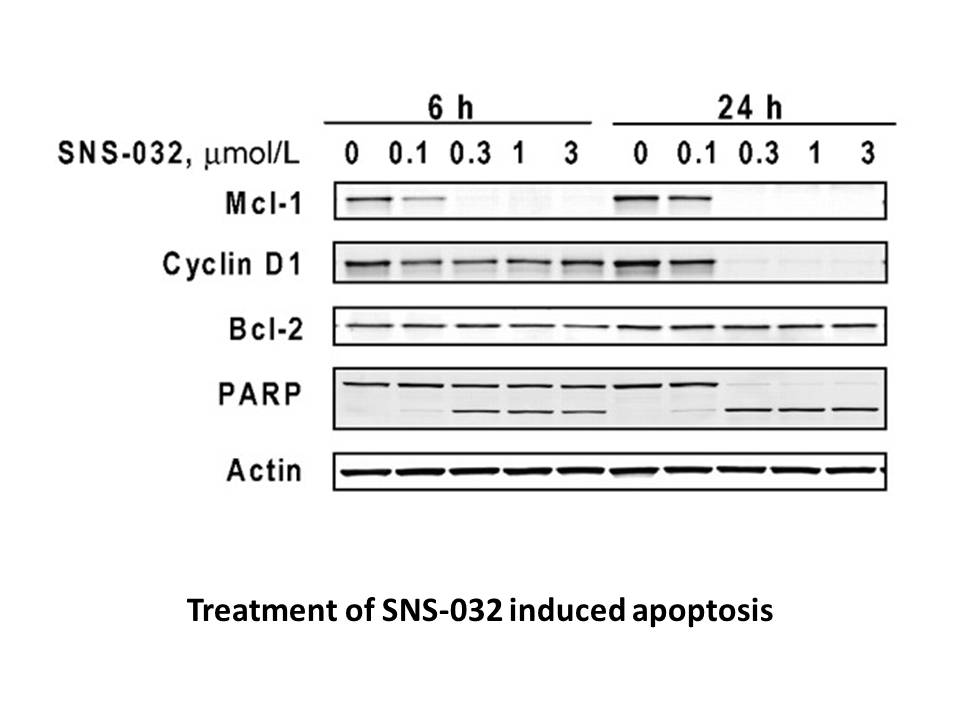
Conclusion Limitations Conflict of interest Introduction The prevalence of KU60019 failure prevalence is increasing throughout the world. The reasons for this pandemic include the aging populations of both industrialized and developing nations and a growing incidence of obesity, diabete
-
Este n mero correspondiente al primer semestre
2019-05-28
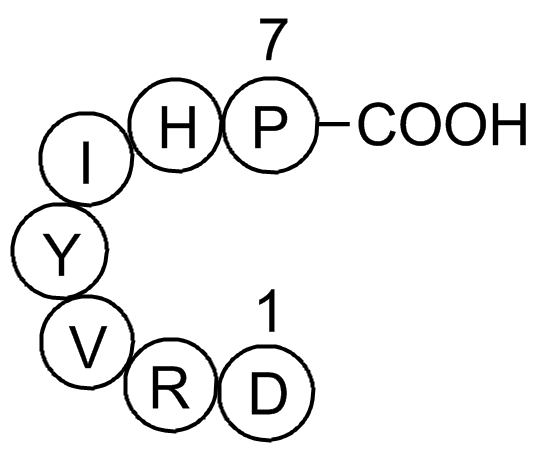
Este número, correspondiente al primer semestre del año 2013, se abre con una sección dedicada treat reflexionar sobre América Latina desde diferentes puntos cardinales: literatura, ciencia y tecnología, derechos humanos, filosofía, historia de las ideas y dos perspectivas del pensamiento de Francis
-
De acuerdo con el historiador argentino la independencia de
2019-05-28
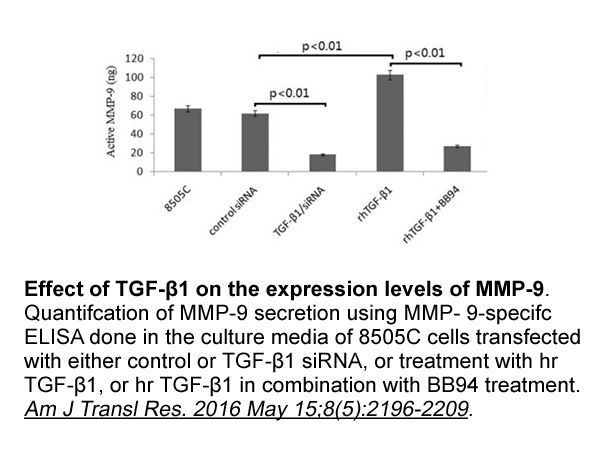
De acuerdo con el historiador argentino, la independencia de las colonias españolas en América es el resultado de una degradación del poder español que había comenzado desde 1795. En primera instancia, el poder de la Corona española se hacía cada vez más lejano, pues la pérdida de la guerra de Españ
-
La no intervenci n en
2019-05-28
La no intervención en los asuntos de otro Estado no significa indiferencia ante los problemas que éste pueda tener. En efecto, algunos usos y gestiones pueden perjudicar 12-O-tetradecanoyl phorbol-13-acetate cost países vecinos y también al mismo Estado que las realiza. Es por esto que la soberanía
-
Es importante subrayar que el recorte del corpus circunscrit
2019-05-28

Es importante subrayar que el recorte del corpus circunscrito al año 1939 también tiene su sustento historiográfico, es sabido que glutamate receptor partir de este momento se establece la figura del exiliado político. Asimismo, terminada la Guerra Civil española y habiéndose decretado la victoria d
16067 records 1028/1072 page Previous Next First page 上5页 10261027102810291030 下5页 Last page